

Embora eu originalmente tenha decidido destacar o trabalho dos pesquisadores do início de carreira no simpósio, a resposta extremamente entusiasta do professor Vasso Episkopou, juntamente com o meu interesse em padronização anteroposterior, fez dela uma escolha óbvia para uma entrevista. No ultimate do primeiro dia, embora eu estivesse cansada de um dia cheio de apresentações interessantes e conversas envolventes, eu me aproximei de seu pôster – que period uma série não convencional, mas eficaz, de slides de powerpoint laminados!
O padrão anteroposterior (AP) em embriões precoces de vertebrados é influenciado pela sinalização nodal – altos níveis promovem destinos anteriores, enquanto níveis baixos acionam destinos posteriores. Portanto, você pode esperar um gradiente de nodal ao longo da sequência primitiva, mas esse não é o caso; É expresso onipresente na sequência. Essa discrepância é o ímpeto da pesquisa realizada por Vasso e seu grupo no Imperial School London.
Snon, um repressor da sinalização nodal, liga o Smad4 para suprimir genes que promovem o destino anterior em elementos de ligação ao Smad (SBES). Após a ativação nodal, o Smad2/3 fosforilado (PSMAD) forma um complexo com snon, que é reconhecido e degradado à sua totalidade pela ubiquitina ligase arkadia (RNF111). Esse mecanismo surpreendente liga diretamente os níveis de PSMAD à redução de SNON, apenas forçando a remoção de SNON sob a alta sinalização do PSMAD. “Os genes alvo anteriores, eles exigem Arkadia e alta sinalização, a fim de degradar o repressor”, explicou Vasso. O SBE agora está claro, permitindo que o PSMAD-SMAD4 se ligasse e, juntamente com os co-ativadores, inicie a transcrição dos genes anteriores. A necessidade de Arkadia para o desenvolvimento anterior é muito clara no fenótipo sem cabeça dos camundongos Arkadia-/-. Além disso, a formação da cabeça é resgatada em embriões Sno-/-; Ark-/-, confirmando que Arkadia é responsável por alcançar a despressão.
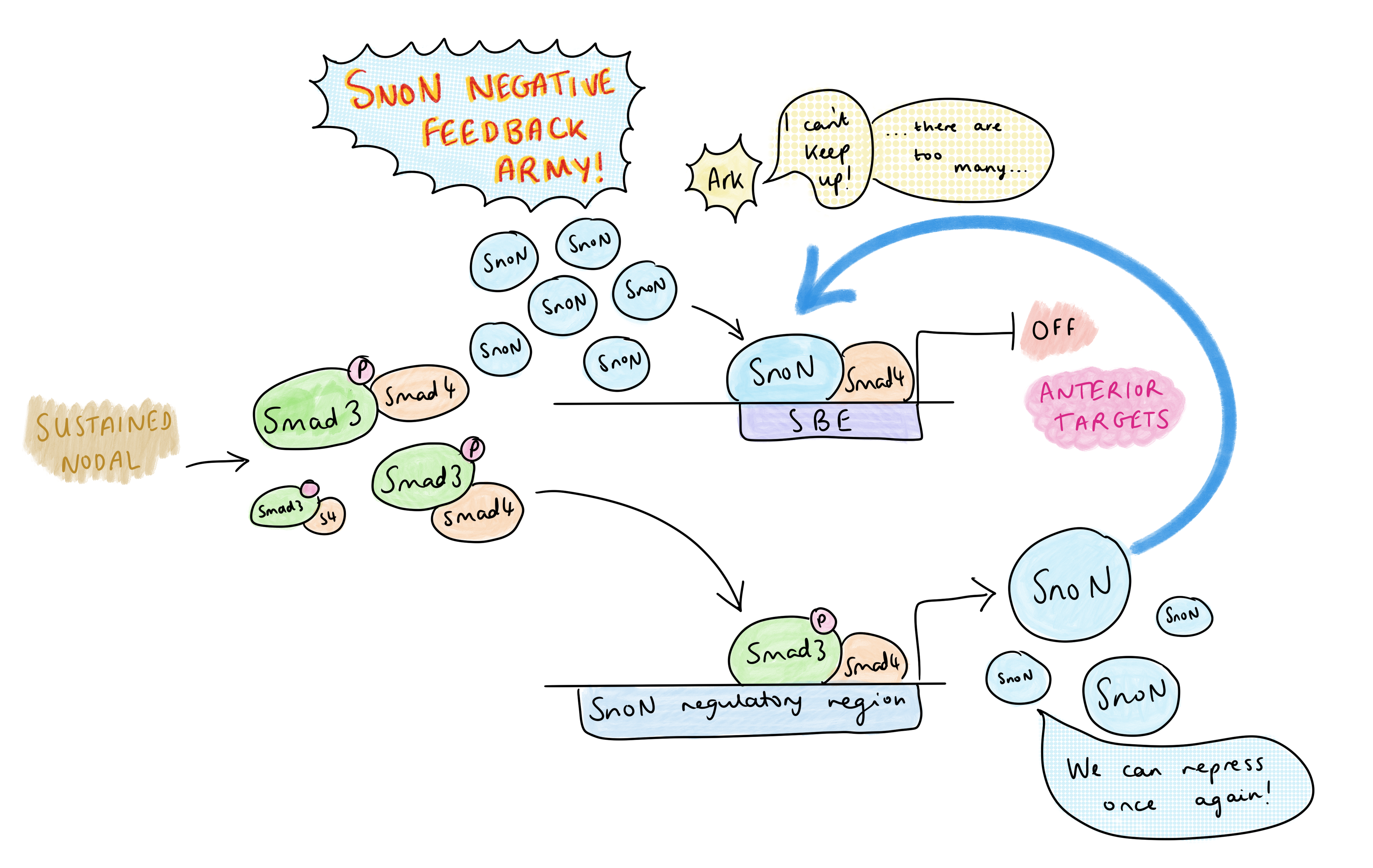
Mas a sinalização nodal é abundante na sequência primitiva – então, como o padrão de AP é regulado? O grupo de pesquisa de Vasso identificou que a dinâmica do padrão de AP de controle de sinalização nodal na faixa primitiva, em vez de gradientes de morfogênio. Um tratamento de curso de tempo TGF-B de células-tronco embrionárias mostrou que, em altos níveis de sinalização, há uma diminuição temporária no SNON em 1-2 horas devido à sua degradação por Arkadia, os níveis de SNON aumentam novamente em 4-6 horas; Essa redução temporária libera espaço no SBE para o PSMAD-SMAD4 para ativar a transcrição dos genes anteriores. O aumento dos níveis de SNON em 4-6H é devido a um efeito de suggestions negativo: um dos principais alvos do PSMAD é o próprio Snon, e o aumento resultante do Snon sobrecarrega Arkadia. Isso leva a uma ativação muito transitória de alvos anteriores anteriores dependentes de Arkadia.
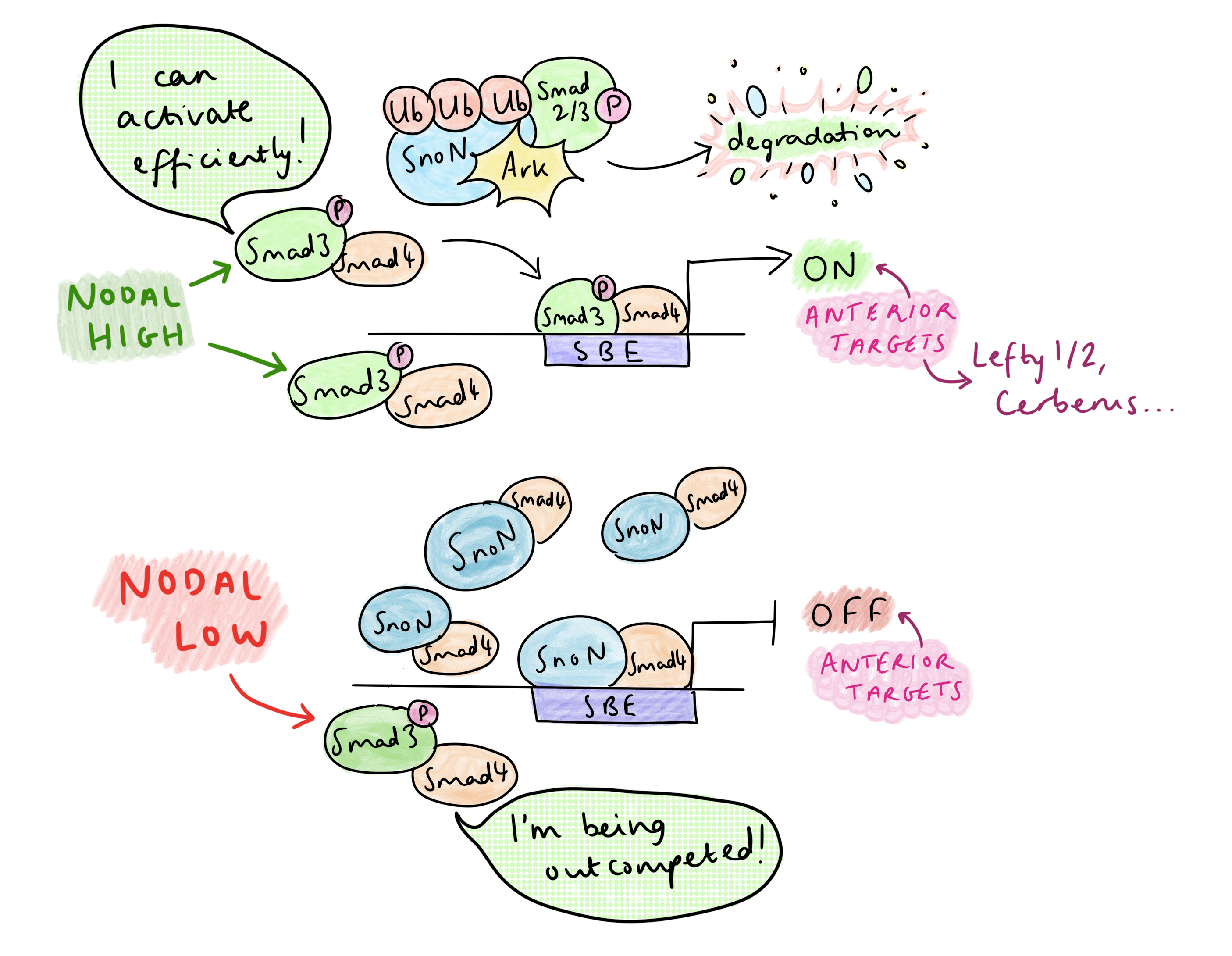
Por outro lado, na presença de níveis mais baixos de nodal, Snon não é degradado por Arkadia; portanto, há concorrência entre Snon-Smad4 e Psmad-Smad4 para ligar o SBE. Os alvos nodais posteriores não exigem que o SBE seja limpo completamente-apenas um baixo nível de sinalização nodal é necessário para relaxar a cromatina, pausando a desacetilação da histona, o que permite que os co-reguladores de alvos posteriores se ligam (em outros locais) e ativem a transcrição. Como os genes posteriores são co-regulados por outros fatores de transcrição, eles precisam apenas de sinalização nodal para que a depressão parcial seja ativada. Isso contrasta com os alvos anteriores, que só podem ser ativados pelo PSMAD-SMAD4 após a limpeza do snon do SBE.
No complete, isso significa que a identidade anterior na faixa primitiva deve ser adquirida em uma janela de tempo muito curta, em resposta a altos níveis agudos de sinalização nodal, antes que qualquer suggestions negativo possa começar. Vasso reuniu isso com o que sabemos sobre a migração celular na sequência – as primeiras células para sair, as células mais rápidas, deixam a tendência. Essas células também expressam antagonistas nodais, como canhoto 1/2 e Cerberus, dobrando para garantir que sua exposição ao nodal tenha vida muito curta. “Cerberus é um ataque triplo”, explicou Vasso, “um inibidor nodal, Wnt e BMP. Portanto, essas células expressam antagonistas a se proteger de sinais sustentados (posteriorizadores) ”. Esses antagonistas ativam a transcrição de genes iniciais imediatos, garantindo que a identidade anterior seja rapidamente adquirida. Por outro lado, as células migratórias mais lentas adquirem identidade posterior, já que permanecendo na sequência por mais tempo as expõe à sinalização nodal sustentada, levando à repressão de genes anteriores e permitindo que os co-reguladores imporem um destino posterior.
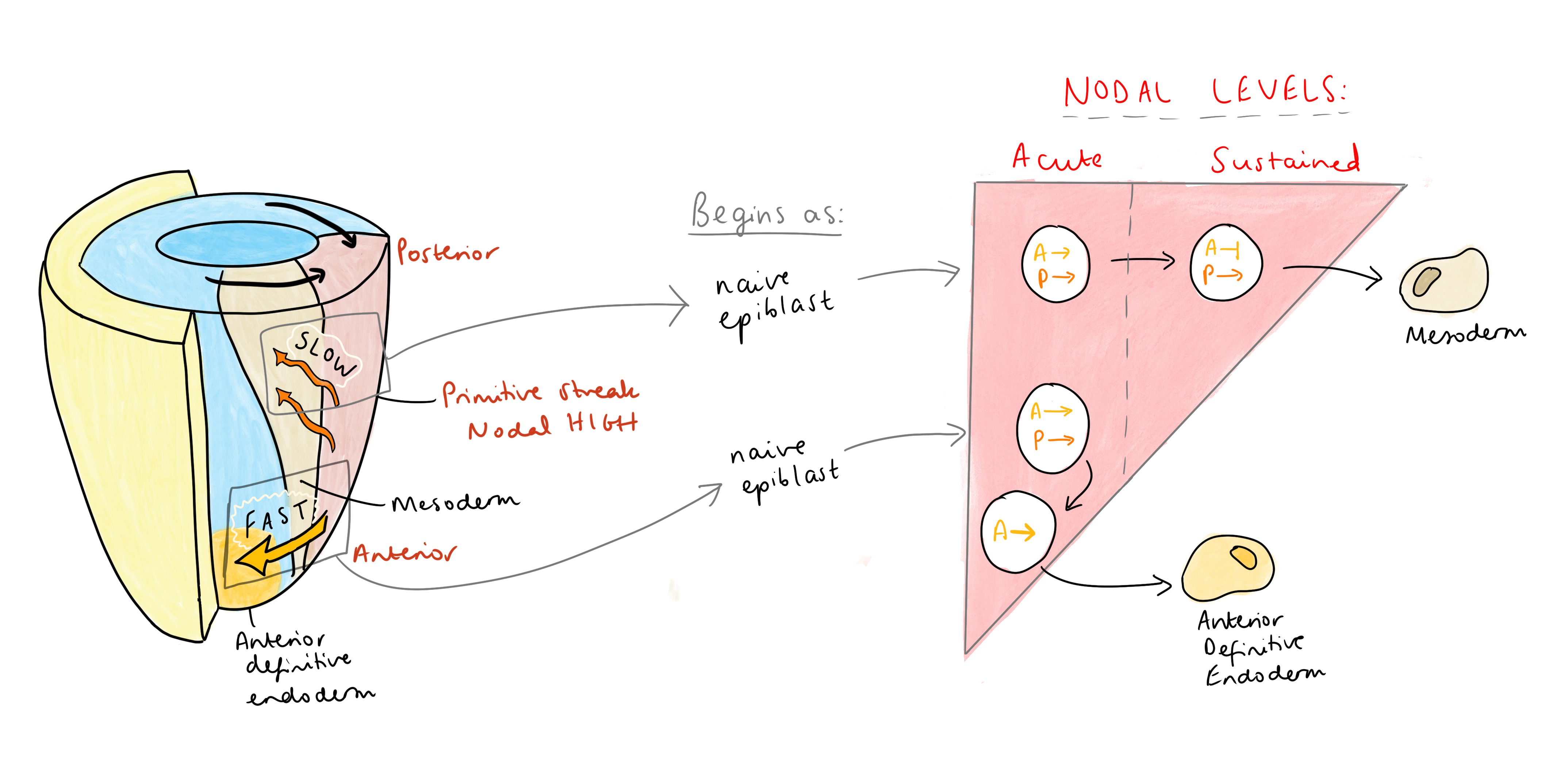
Vasso sugere que essa regulação dinâmica da sinalização pode ser um mecanismo generalizado na determinação do destino. Um princípio semelhante se aplica à diferenciação de linfócitos T: os altos níveis de TGFβ impulsionam a diferenciação de Treg de maneira dependente de Arkadia, enquanto níveis baixos impulsionam a diferenciação Th17 independentemente de Arkadia (Xu et al.2021). Este modelo pode se estender a outros contextos biológicos ainda a serem explorados.
Você pode ler a pré -impressão do grupo de Vasso sobre a dinâmica nodal aqui:
Carthy, JM, Ioannou, M. e Episkopou, V. (2019) ‘Arkadia by way of Snon permite que efetores de sinalização Nodal-Smad2/3 transcrevam genes diferentes, dependendo de seus níveis’. Biorxiv, p. 487371. Disponível em: https://doi.org/10.1101/487371.
Leia sobre como o mesmo sistema age na diferenciação de linfócitos T aqui:
Xu, H. et al. (2021) ‘A sinalização de Arkadia-Ski/Snon regula diferencialmente a diferenciação de células ITREG e Th17 induzida por TGF-β’, Jornal de Medicina Experimental218 (11), p. E20210777. Disponível em: https://doi.org/10.1084/jem.20210777.
Fique ligado para mais entrevistas de pôsteres em breve!
O submit 3º Simpósio de Crick Beddington: destinos anteriores esperam aqueles que migram – como a dinâmica nodal molda o padrão anteroposterior apareceu primeiro o nó.




")

{kind=link}