

Visão geral
Em quatro epitélios de camada de murina, a primeira transição arquitetônica 3D – internalização de células durante a formação de placode – trata a quebra de simetria. Yap lê a posição da célula; Notch comete vizinhos para destinos basais vs luminais1.
Como o projeto começou
Este projeto começou com uma simples observação inspirada em pesquisas em nosso laboratório sobre o desenvolvimento da glândula mamária em estágio inicial2: que mesmo muito cedo no desenvolvimento, quando o tecido ainda não começou a ramificar a morfogênese, o destino celular já está espacialmente organizado na glândula mamarsa embrionária. Esse padronização de tecido segregando linhagens celulares distintas pareciam muito robustas e nos perguntamos quais vias estavam envolvidas em tais decisões do destino inicial.
Por que esses tecidos?
Optamos por estudar as glândulas mamárias, lacrimais, salivares e próstata por três razões:
- Eles compartilham uma arquitetura em camada e a mesma hierarquia celular com células -tronco, dando origem a células basais e luminais,
- Durante a especificação do destino, eles mostram assinaturas transcricionais semelhantes e dinâmica da restrição de potência do destino e
- Todos eles são epitélios ramificados.
Em outras palavras, diferentes órgãos com origens embrionárias distintas, mas uma lógica estrutural comum, portanto, o paradigma experimental perfeito para testar se um mecanismo conservado está subjacente à compartimentalização do tecido precoce e à segregação do destino.
O que aprendemos
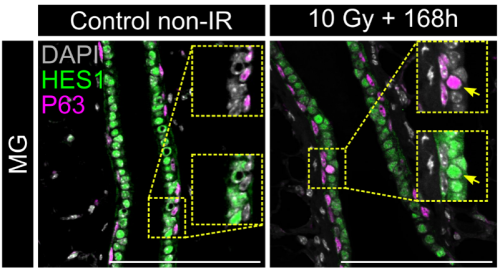
Nos organoides e explantes de tecidos embrionários, a quebra de simetria coincidiu com a internalização das células: as células internas adquirem alta atividade de Notch (Hes1) enquanto as células externas retêm a expressão de p63 e o YAP nuclear (Figura 1). O YAP atua como o intérprete de posição – é uniforme em todas as células antes da compartimentação de internalização/tecido, enquanto se restringe espacialmente posteriormente. Por outro lado, o Notch atua como maquinaria de compromisso, necessário e suficiente para impulsionar a identidade das células luminais. Perturbações que reforçam a atividade YAP uniformes mantêm as células em um estado híbrido p63⁺/hes1⁺ e atrasam a compartimentalização do tecido, enquanto a ativação de Notch substitui esse bloqueio na diferenciação e impõe a aquisição de destino luminal.
Na regeneração de tecidos adultos (induzida por ablação ou irradiação de células luminais), observamos as mesmas células híbridas p63⁺/hes1⁺ e um aumento de células que abrigam YAP nuclear, reavalindo o estado pré-comprometido no desenvolvimento precoce. O tamanho crítico do tecido no qual a quebra de simetria ocorre é maior in vivo do que nos organoides, provavelmente porque as entradas de nicho modulam a sinalização YAP, atrasando o comprometimento celular, apesar da geometria semelhante.
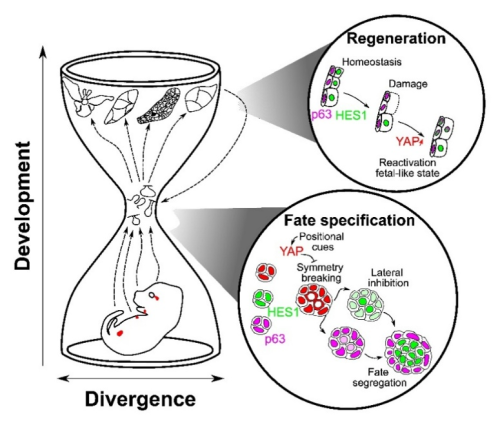
Uma lógica conservada “ampulheta”
Apesar das origens distintas (glândulas exócrinas derivadas de ectoderme vs. próstata derivada de endoderme), esses órgãos parecem reutilizar um equipment de ferramentas comum no ponto de quebra de simetria e comprometimento das células-tronco. Diversas entradas a montante, incluindo geometria, nicho e mecânica de tecidos, convergem para o YAP no gargalo de uma ampulheta, que depois as interações de portas Notch -P63 para resolver o destino. Mais tarde, no desenvolvimento, os tecidos divergem novamente seguindo programas específicos de órgãos adaptados às diferentes funções de cada tecido. A alta conservação deste gargalo médio, YAP → Notch/P63, é o que dá ao mecanismo tanto a robustez quanto a portabilidade em diferentes contextos, incluindo a regeneração.
Levar para casa
A arquitetura de tecidos inicia, o YAP interpreta, o Notch resolve. A internalização de células atua como a sugestão determinística que converte a forma do tecido em destino celular entre os epitélios de camada em bicamada – e a mesma lógica é reimplantada durante o reparo.
1. REVISTÃO, RP, HUYGHE, M., Barthelemy, A., Couto-Moreira, H., Deshayes, T., Harari, L., Sumbal, J., Faraldo, MM, Dubail, M., Fouillade, C., et al. (2025). Os sinais conservados controlam a auto-organização e a quebra de simetria dos epitélios de camadas murinos durante o desenvolvimento e regeneração. Dev. Célula. https://doi.org/10.1016/j.devcel.2025.06.007.
2. Carabaña, C., Solar, W., Veludo Ramos, C., Huyghe, M., Perkins, M., Maillot, A., Rediot, R., Hartani, F., Faraldo, MM, Lloyd-Lewis, B., et al. (2024). Espacialmente, subconjuntos de células epiteliais e mesenquimais distintas ao longo da restrição de linhagem progressiva na glândula mamária embrionária ramificada. EMBO J., 1–29. https://doi.org/10.1038/S44318-024-00115-3.
O put up Geometria Primeira: como as pistas posicionais ditam o destino em epitélios em bicamada apareceu primeiro o nó.






{kind=link}