

Escrito por Isaia Vardanega
Atrás da história do papel de “A sinalização de clavata formas”.
As gramíneas desempenham um papel essential na produção de alimentos humanos e são cultivados principalmente por seus grãos comestíveis. A quantidade de grãos que uma espécie de grama pode produzir está intimamente ligada à arquitetura de sua inflorescência. Ao longo da evolução, as gramíneas desenvolveram uma ampla variedade de arquiteturas de inflorescência. Das inflorescências ramificadas complexas do Oryzae Tribo (por exemplo, arroz), onde vários grãos se desenvolvem em ramos primário e secundário, para a inflorescência simples do tipo pico da Triticeae Tribo (por exemplo, cevada e trigo), onde grãos únicos se desenvolvem em eixos vestigiais curtos chamados Rachillae1 (Fig.1 a).
Essas diferenças arquitetônicas são estabelecidas durante os estágios iniciais do desenvolvimento da planta pela atividade de meristemas, estruturas especializadas que lideram o crescimento dos órgãos. O tamanho, a posição e a vida útil desses meristemas determinam a eventual forma da inflorescência2. Um exemplo de como pequenas diferenças na atividade do meristema podem afetar significativamente a arquitetura remaining da inflorescência é encontrada dentro do Triticeae tribo. Na cevada, o Rachilla Primordium cresce apenas o suficiente para formar uma única florzinha e grãos. Por outro lado, em trigo, sua atividade prolongada permite a formação de múltiplas florzinhas por espiguetas, resultando em sua característica espizagem de vários grãos1 (Fig.1 b, c).
Figura 1: (A) Representação esquemática de diferentes arquiteturas de inflorescência de grama. Principais caule e galhos em verde escuro, grãos em verde claro e Rachillae em vermelho. (B, c) Imagens de microscópio eletrônico de varredura de inflorescências de cevada e trigo em estágios iniciais de desenvolvimento. Diferentes órgãos compreendendo um espiguetista são coloridos (Rachilla Primordium em vermelho, meristema de floret em amarelo e lema primordium em verde). Cada espiguetista se desenvolve em um único grão na cevada e vários grãos de trigo.
Quando entrei para o laboratório de Rüdiger Simon para iniciar meu doutorado, o grupo estava focado principalmente em entender o papel das vias de sinalização da CLE na regulação da forma, tamanho e manutenção de meristemas apicais de tiro e raiz em Arabidopsis Thaliana. Naquela época, eles começaram a estender suas pesquisas à cevada de plantas de cereais. Gwendolyn Kirschner, ex-aluno de doutorado no laboratório, começou a investigar o papel da sinalização do cle-peptídeo na cevada, gerando linhas de repórteres fluorescentes, incluindo os ortólogos da cevada do Arabidopsis Peptídeo CLE40 (HVFCP1) e CLV1 receptor (Hvclv1), que anteriormente demonstrou common o destino das células -tronco em Arabidopsis meristemas3,4. Enquanto Gwendolyn analisou principalmente as raízes de cevada, meu projeto se concentrou mais no meristema apical e no desenvolvimento de inflorescência.
Em comparação com a simples inflorescência de Arabidopsisas gramíneas evoluíram uma organização mais complexa, com diferentes tipos de meristemas levando à formação de vários órgãos que compreendem o espiguetista, a unidade básica responsável pelo desenvolvimento de grãos em cereais. Essa observação nos levou às perguntas: “Como a forma e a atividade de todos esses meristemas diferentes são regulados e coordenados para gerar inflorescências específicas em gramíneas? A cevada evoluiu as vias de sinalização CLE/clavata específicas para common a atividade de diferentes tipos de meristemas?”
O HVCLV1 regula a atividade do meristema ao longo dos eixos verticais e laterais da inflorescência da cevada.
Comecei mutando o ortólogo mais próximo de CLV1 em cevada (Hvclv1) e outros receptores intimamente relacionados, que ainda estamos investigando. Em milho, como em Arabidopsismutação de CLV1 ou seu ortólogo leva a um meristema de inflorescência drasticamente aumentada, resultando na formação desorganizada de primórdios adicionais4,5. Na cevada Hvclv1 Mutante, inicialmente observei a formação ocasional de grãos extras dentro da inflorescência. No entanto, a análise detalhada da microscopia eletrônica de varredura revelou que um meristema de inflorescência aumentado gerou uma linha adicional de primordia espiguatária, organizada em uma filotaxia em espiral e não de maneira desorganizada (Fig. 2a, b).
Além disso, eu notei que Hvclv1 As inflorescências desenvolveram um Rachilla Primordium alongado, que produziu florzinhas adicionais por espiguetas, um efeito observado anteriormente em mutantes de cevada como multiflorus2.b e intermediário-m6,7. Esses resultados me levaram a concluir que os grãos ectópicos gerados pelo Hvclv1 Os mutantes foram devidos ao aumento da atividade do meristema de inflorescência ao longo do eixo vertical e do primordio de Rachilla ao longo do eixo lateral. Para apoiar ainda mais, fotografi Rachillae madura da WT e Hvclv1 plantas e observou a formação de uma estrutura semelhante a meristema na ponta do Hvclv1 Rachilla, que se transformou em um ramo pequeno de crescimento ativamente, em vez da estrutura peluda vestigial observada no WT (Fig. 2C, D).

Figura 2: (A, b) Imagens de microscópio eletrônico de varredura exibindo filotaxia de espiguetas (linhas tracejadas) em WT (a) e Hvclv1 (B) Dicas de inflorescência precoce, combinadas com fotos das respectivas inflorescências finais. (C, d) Imagens de microscópio eletrônico de varredura de WT e Hvclv1 Inflorescências iniciais. As cores foram usadas para destacar diferentes meristemas e primórdios (meristema de inflorescência em azul, meristema de espigina no violeta, rachilla primordium em vermelho, meristema de floret em amarelo, antera e carpes primordia em marrom e rosa). Linhas tracejadas vermelhas indicam a rachilla madura, combinada com imagens de zoom onde a seta branca indica a estrutura do tipo meristema identificada em Hvclv1. Figura modificada de Vardanega et. Al 2025.
O peptídeo CLE HVFCP1 Atos com Hvclv1 para restringir a atividade do Primordium Rachilla à formação de um único florzato.
Uma vez que a função do Hvclv1 O receptor foi caracterizado, eu me perguntava se a regulação do crescimento de Rachilla foi determinada pela ligação de um peptídeo CLE específico. A cevada possui 28 peptídeos CLE diferentes, mas eu estava procurando especificamente um que é fortemente conservado entre as gramíneas e pode ter contribuído para a redução drástica do tamanho da filial em Triticeae. Ao revisar a literatura, percebi que apenas um peptídeo mantinha a mesma sequência de proteínas em todas as espécies estudadas de grama: FCP18. HVFCP1 é o ortólogo mais próximo da Arabidopsis CLE40para o qual já tínhamos uma linha de repórter fluorescente. Quando eu examinei a expressão de HVFCP1 Durante o desenvolvimento da inflorescência de cevada, notei que não foi apenas co-expresso com Hvclv1 mas também especificamente expresso no Primordium Rachilla (Fig. 3 A, B). A insensibilidade do Hvclv1 atirar meristema apical para HVFCP1 tratamento peptídico, juntamente com a formação de uma Rachilla alongada produzindo floretes adicionais, mesmo em HVFCP1 mutantes (Fig. 3 c), finalmente demonstraram que HVFCP1 interage com Hvclv1 common a atividade de Rachilla na cevada, determinando assim sua arquitetura de inflorescência específica.
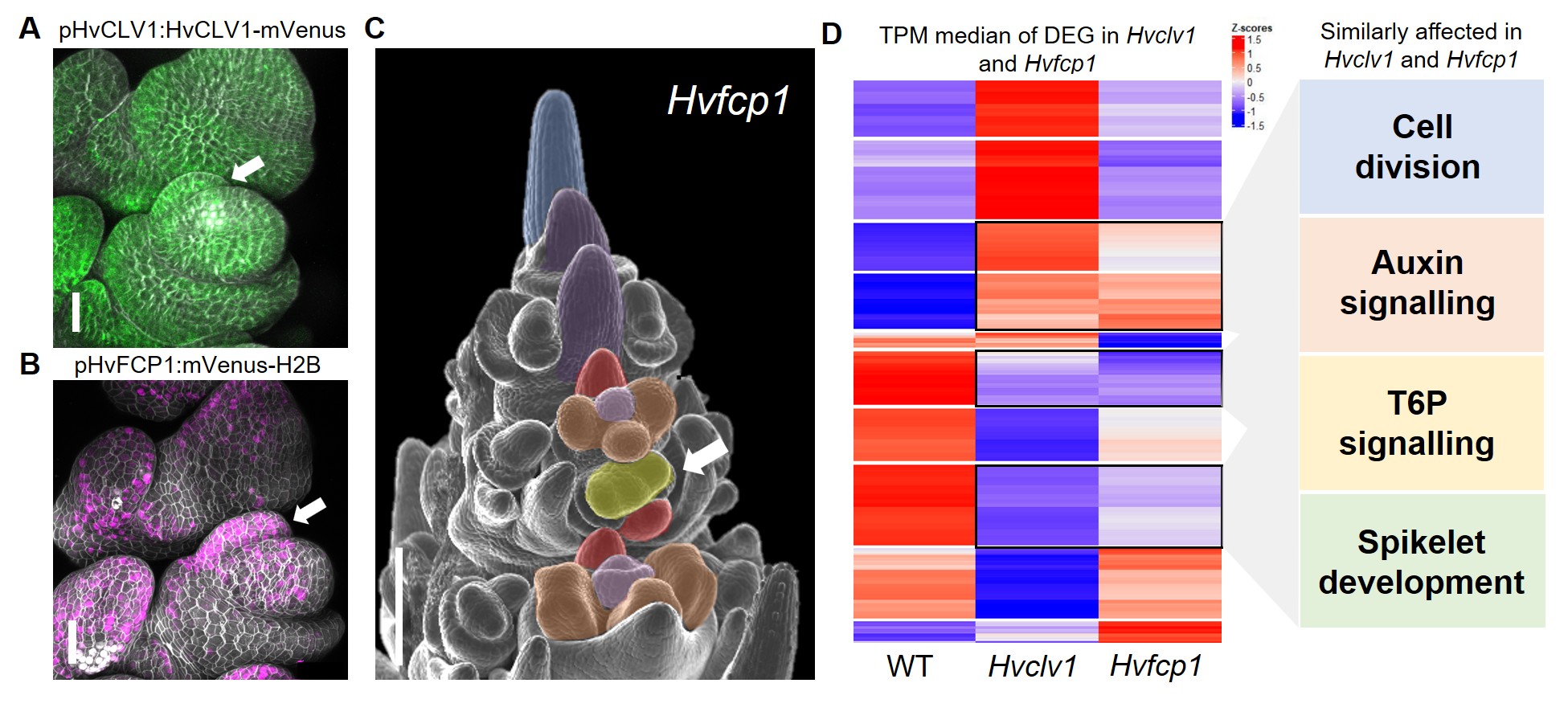
Figura 3: (A, b) Imagens confocais de microscópio de espiguetas de cevada exibindo Hvclv1 Localização de proteínas (verde) e HVFCP1 Atividade do promotor (magenta). A seta branca indica o Primordium Rachilla. (C) HVFCP1 Inflorescência exibindo florzinhas duplas (seta branca). (D) Resultados do RNA seq. O mapa de calor mostra o escore z de transcrições medianas por milhão (TPM) valores para cada um dos genes expressos de maneira diferente (deg) em Hvclv1 vs wt e HVFCP1 vs wt. Retângulos pretos indicam genes similarmente expressos diferencialmente Hvclv1 vs wt e HVFCP1 vs WT, com os processos biológicos afetados ao lado. Figura modificada de Vardanega et. Al 2025.
Investigei então quais genes são direta ou indiretamente regulados por HVFCP1/Hvclv1 realizando sequenciamento de RNA em inflorescências mutantes em comparação com o tipo selvagem (WT). A análise do transcriptoma revelou vários genes similarmente expressos diferencialmente (deg) entre Hvclv1 vs wt e HVFCP1 Vs WT, envolvido em processos como divisão celular, sinalização de auxina e sinalização de trealose-6-fosfato, fornecendo possíveis genes-alvo que estamos investigando atualmente (Fig.3 D).
Traduzir conhecimento e técnicas de plantas modelo para culturas.
Além de sua novidade biológica, este artigo representa um exemplo bem -sucedido de pesquisa translacional, técnicas de ponte e conhecimento de espécies modelo para culturas agronomicamente significativas. Aplicamos várias técnicas de microscopia mais comumente usadas em plantas modelo, como Arabidopsis Thalianamas com menos frequência empregada em biologia de cereais. Desenvolvemos linhas de repórter e complementação e quantificou a internalização citoplasmática do receptor em meristemas de Rachilla e Floret. Além disso, utilizamos métodos como reconstrução 3D e smRNA-Fish para análise fenotípica detalhada do meristema de inflorescência e seus padrões de expressão.
Para desenvolver essa abordagem, geramos Barvista (http://purl.org/barvista/house), um conjunto de dados que fornece informações transcricionais para cada célula dentro da inflorescência da cevada, integrando dados de sequenciamento de RNA de célula única com resultados transcriptômicos espaciais. Usando esse recurso, identificamos fatores de transcrição envolvidos no estabelecimento dos padrões específicos de ontogênese do meristema necessários para moldar a morfologia característica do pico de cevada9.
Em conclusão, nosso trabalho esclarece as vias de sinalização que regulam a forma e o comportamento dos tipos individuais de meristemas dentro da inflorescência, abrindo caminho para esforços futuros para projetar a arquitetura de inflorescência por meio da regulação direcionada de atividades distintas de meristemas.
Referências:
- Koppolu, R. & Schnurbusch, T. Caminhos de desenvolvimento para moldar a arquitetura de inflorescência de pico em cevada e trigo. Journal of Integrative Plant Biology 61278-295 (2019).
- Kyozuka, J., Tokunaga, H. & Yoshida, A. Controle da inflorescência da grama Formam pelo ajuste fino da mudança de fase do meristema. Opinião atual em biologia vegetal 17110-115 (2014).
- Berckmans, B., Kirschner, G., Gerlitz, N., Stadler, R. & Simon, R. Cle40 A sinalização regula o destino das células -tronco da raiz. Plant Physiol 1821776-1792 (2020).
- Schlegel, J. et al. Controle da homeostase das células -tronco de Arabidopsis por duas vias antagônicas de sinalização de peptídeos CLE. eLife 10E70934 (2021).
- Bommert, P. et al. O anão de borla espesso codifica um ortólogo putativo de milho da reincidência rica em leucina de Arabidopsis Clavata1 quinase do tipo receptor. Desenvolvimento 1321235-1245 (2005).
- Koppolu, R. et al. O mutante de cevada multiflorus2.b revela variação genética quantitativa para a nova arquitetura de espiguetas. Theor Appl Genet 135571-590 (2022).
- Zhong, J. et al. O intermediário-M codifica um ortólogo HVAP2L-H5 e é necessário para a inflorescência indeterminação e a determinação de espiguetas na cevada. Anais da Academia Nacional de Ciências 118e2011779118 (2021).
- Goad, DM, Zhu, C. e Kellogg, EA Identificação e agrupamento abrangente de genes relacionados a CLV3/ESR (CLE) em plantas encontram grupos com função potencialmente compartilhada. Novo fitologista 216605-616 (2017).
- Demesa-Arevalo, E. et al. A imputação integra dados de expressão de genes de célula única e espacial para resolver redes transcricionais no desenvolvimento do meristema de brotos de cevada. 2025.05.09.653223 Pré -impressão em Biorxiv https://doi.org/10.1101/2025.05.09.653223 (2025).
O publish Da ponta ao grão: esculpindo a inflorescência de cevada através da regulação da atividade do meristema apareceu primeiro o nó.






{kind=link}