

Mecanismo da reação Masamune-Bergman. Parte 3: O estado de transição para modelos de Calicheamicina.
Caliqueamicina foi apontado no publish anterior como um produto pure com propriedades antitumorais e com muitas características estruturais estranhas, como um motivo “enedidyne” incomum. A representação é mostrada abaixo.
Uma estrutura parcial mostrada abaixo para Calicheamicina substitui o -(CH2)4- subestrutura com uma cadeia de quatro carbonos que inclui dois sp2centros em vez de dois sp3 centros. O objetivo é descobrir como essas modificações estruturais no clássico Bergman afetam o mecanismo.
TS1 é mostrado abaixo para este modelo e a barreira de energia livre calculada para esta ciclização é de 42,5 kcal/mol no nível uωB97XD/Def2-TZVPP, 2> = 0,345. Dados FAIR DOI: 10.14469/hpc/14583(citar)10.14469/hpc/14583(/citar). Isto se compara com 33,0 kcal/mol calculado para o -(CH2)4- versão, para a qual 2> = 0,266. Para se preparar para modelar o completo Caliqueamicina molécula, a base definida para este modelo foi reduzida a Def2-SVPP e neste nível ΔG‡ foi de 43,0 kcal/mol, 2> = 0,368, sendo a diferença suficientemente pequena para que a redução no conjunto de bases pareça pouco provável que afecte os resultados. Os comprimentos de formação da ligação CC são 1,957 (Deft-TZVPP) e 1,989Å (Def2-SVPP).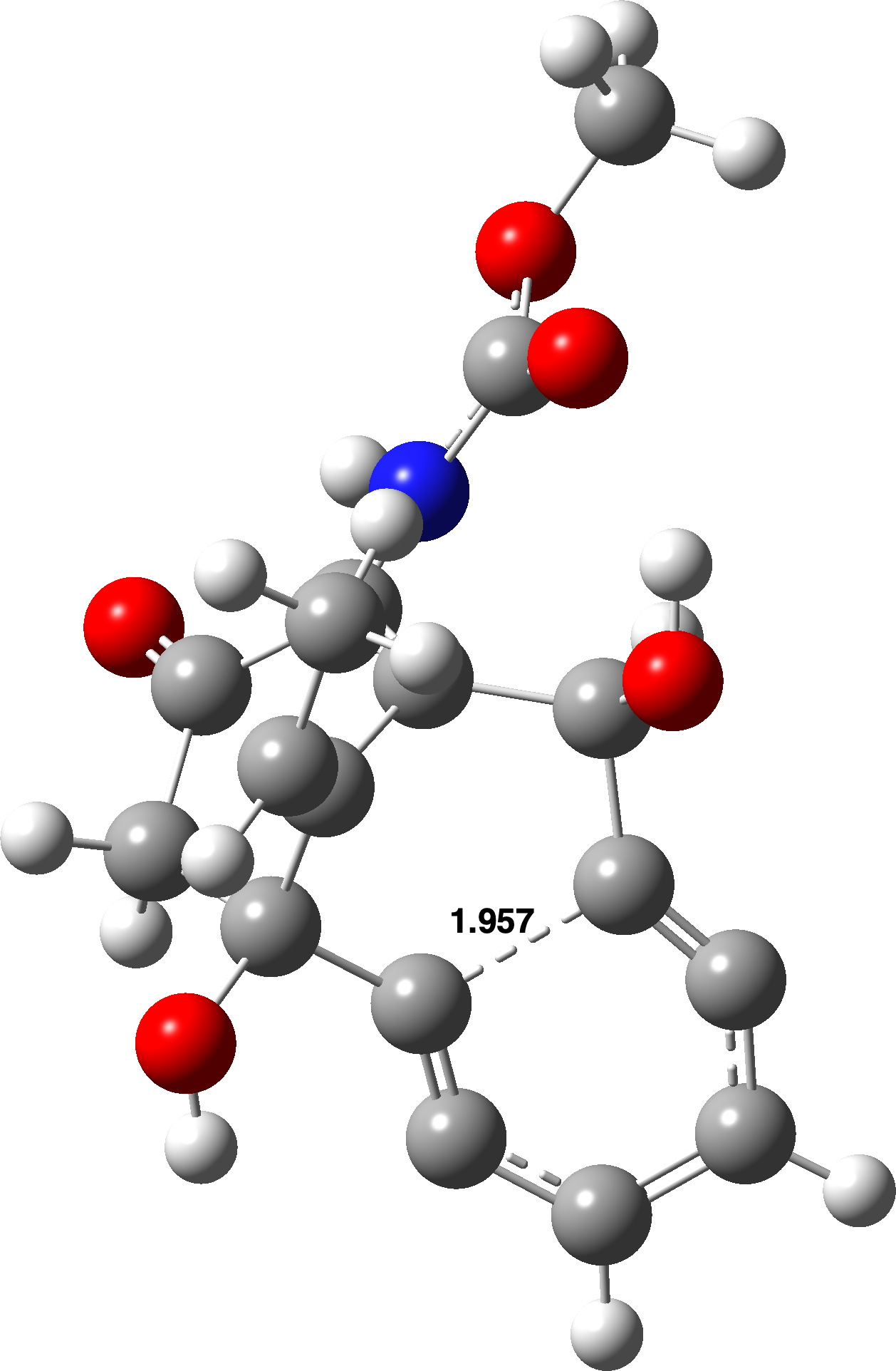
Agora, para um modelo maior contendo toda a molécula de Calicheamicina. Duas possibilidades foram exploradas; aquele em que a geometria do sistema foi totalmente otimizada isoladamente para produzir uma conformação para Calicheamicina que se dobrou sobre si mesma e para a qual ΔG‡ (Def2-SVPP) 40,1 kcal/mol, 2> 0,368.
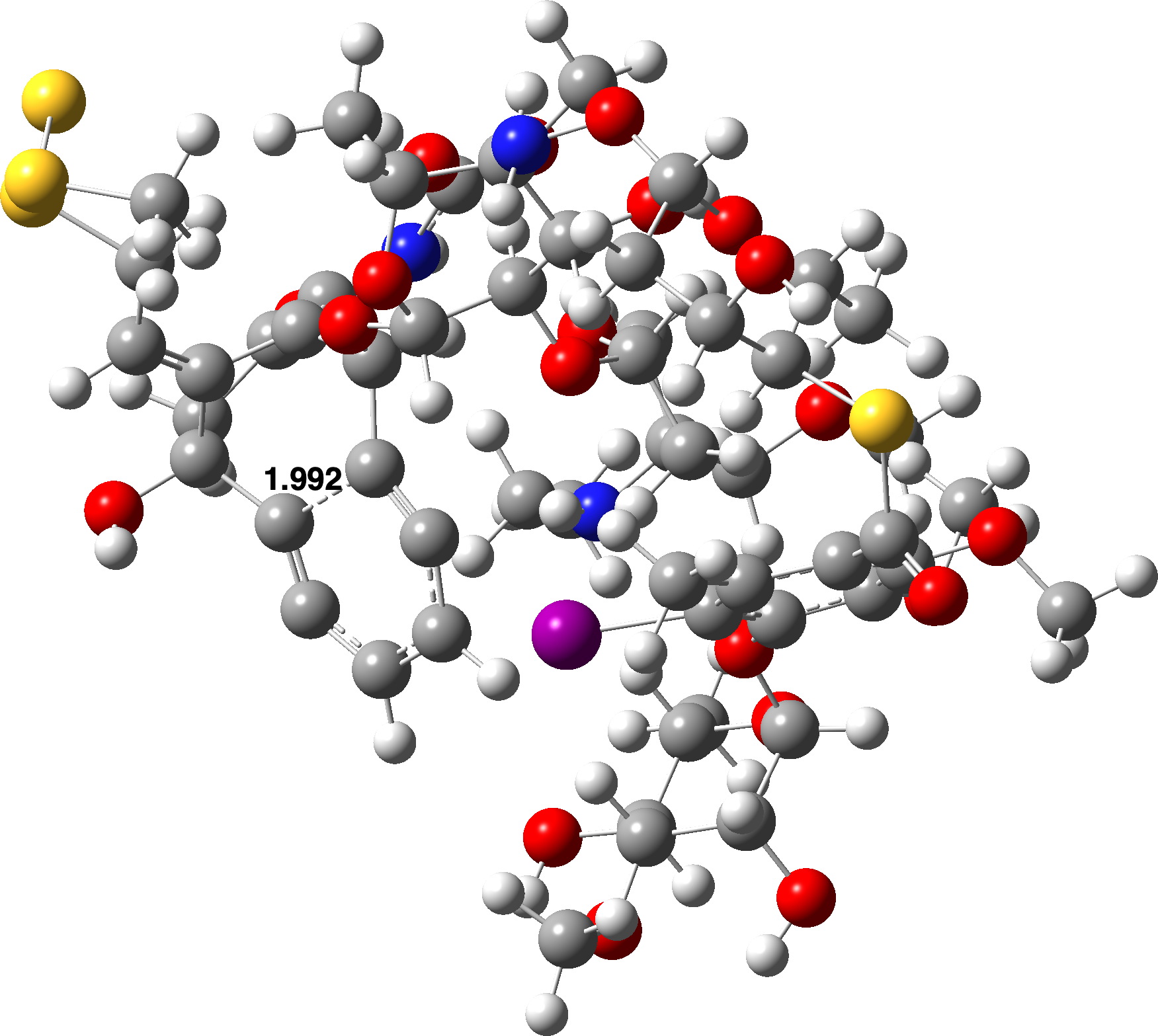
O segundo modelo utilizou a geometria inicial da Calicheamicina obtida a partir de uma estrutura cristalina do ligante dobrada no bosque menor de um fragmento de DNA e que possui uma forma muito mais linear. O reagente neste modo foi +6,1 kcal/mol mais alto em energia do que o anterior e TS1 foi 4,6 kcal/mol mais alto, levando a ΔG‡ 38,6 kcal/mol, 2> 0,367.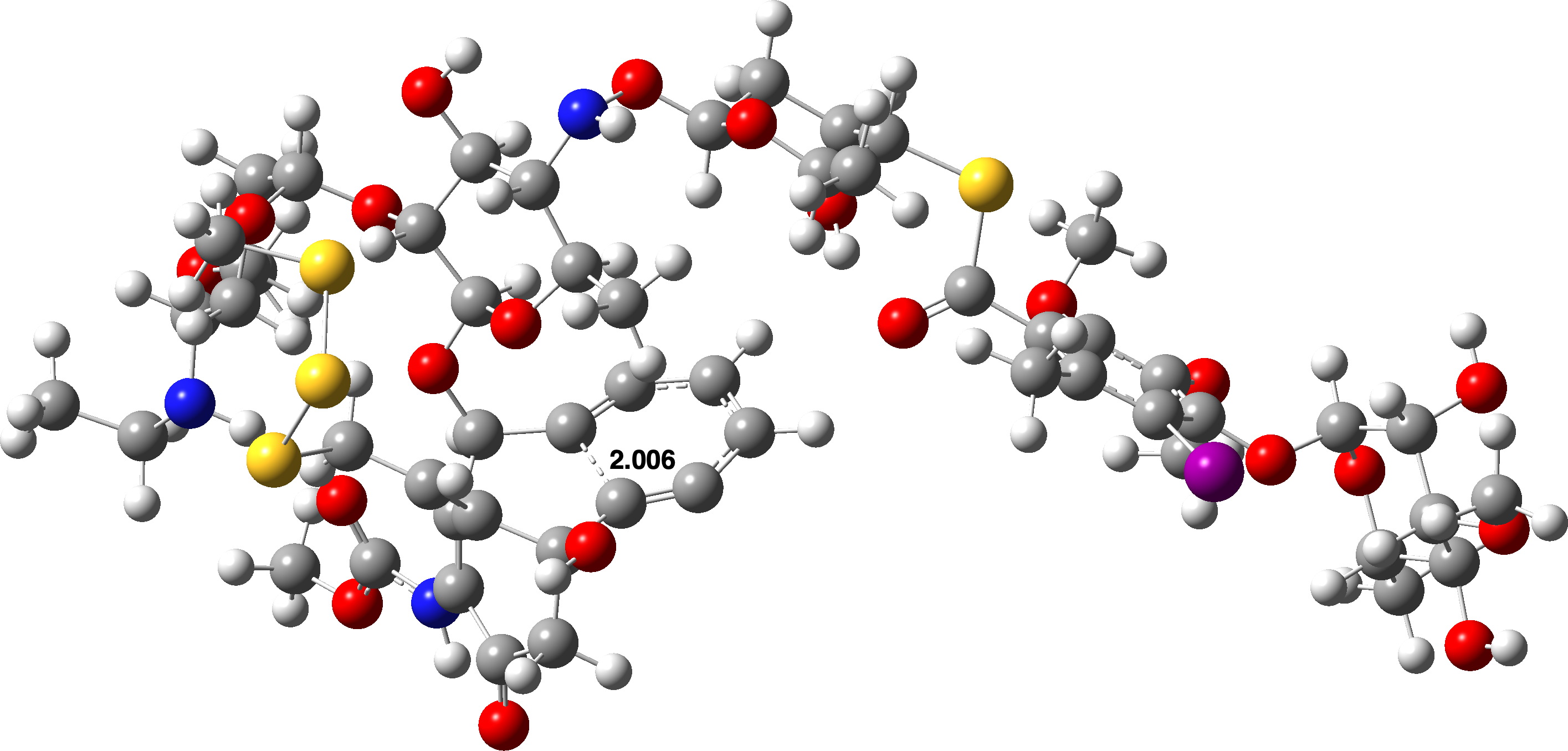
Então, que conclusões podemos tirar destes modelos alargados da ciclização de Bergman? As energias livres de ativação para todos os três modelos estão na faixa de 42,5 – 38,6 kcal/mol, o que é muito superior a um valor compatível com uma reação fácil à temperatura ambiente (~22±3). A observação de que a caliqueamicina pode de facto ser caracterizada como uma estrutura cristalina quando ligada ao ADN sugere que a barreira de ciclização não pode ser demasiado baixa, mas, inversamente, o intervalo de 42,5 – 38,6 kcal/mol parece demasiado grande para que a caliqueamicina seja facilmente activada num biradical, a fim de para abstrair o átomo de hidrogênio e acabar causando a cisão da fita. Poderia o modelo simplista de uma função de onda UHF dividida resultando em valores de 2> 0,37 é o problema? Bem, uma abordagem semelhante foi adotada para modelar o rearranjo de Stevens (cite)10.59350/4010f-fvr26(/cite). Usando uma função de onda de concha fechada simples não biradical, uma barreira de ~48 kcal/mol foi obtida, mas esta foi reduzida para 14 kcal/mol quando o método UHF foi aplicado (2> 0,421), portanto este modelo parece funcionar bem nessas circunstâncias. O júri ainda deve decidir se o mecanismo de ciclização de Bergman está a ser correctamente modelado aqui ou se algo mais complexo está a acontecer.
Relacionado
Esta entrada foi publicada quarta-feira, 11 de setembro de 2024 às 12h22 e arquivada em Química interessante. Você pode acompanhar quaisquer respostas a esta entrada através do RSS 2.0 alimentar. Você pode deixe uma respostaou trackback do seu próprio web site.




